The special creationist claim that there are no transitional fossils is false, as even a cursory search of the internet will demonstrate. The evidence for large-scale evolutionary change as seen in the fossil record is simply beyond dispute. For example, Kathleen Hunt's well-received article on transitional fossils covers only vertebrate fossils, and as it was written in 1997 does not incorporate stunning discoveries such as the 'fishapod' Tiktaalik roseae. Despite this, as she concluded:
Creationists often state categorically that "there are no transitional fossils". As this FAQ shows, this is simply not true. That is the main point of this FAQ. There are abundant transitional fossils of both the "chain of genera" type and the "species-to-species transition" type. There are documented speciations that cross genus lines and family lines. The interpretation of that fact I leave up to you. I have outlined five possible models above, and have explained why I think some of them are better than others. You might disagree with my conclusions, and you can choose the one you think is best, (or even develop another one). But you cannot simply say that there are no transitional fossils, because there are.
As Gould said (1994): "The supposed lack of intermediary forms in the fossil record remains the fundamental canard of current antievolutionists. Such transitional forms are scarce, to be sure, and for two sets of reasons - geological (the gappiness of the fossil record) and biological (the episodic nature of evolutionary change, including patterns of punctuated equilibrium and transition within small populations of limited geological extenet). But paleontologists have discovered several superb examples of intermediary forms and sequences, more than enough to convince any fair-minded skeptic about the reality of life's physical genealogy." [1]
Covering the evidence in full is beyond the scope of this website, but a few posts covering major transitions will suffice to show the vacuity of the special creationist claim that there are no transitional fossils. This post will cover the evidence detailing tetrapod evolution from lobe-finned fish. [2]
To fully grasp the significance of the transitional fossils demonstrating the evolution of tetrapods, let's look at the phylogenetic tree in a little more detail. Fish and the other vertebrates with jaws (amphibians, reptiles, birds, mammals) form a clade known as the gnathostomata. The gnathostomata and lampreys form a group known as craniates.
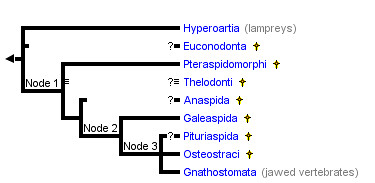
Source: http://tolweb.org/Vertebrata/14829
(*) denotes extinct group.
(?) denotes uncertain phylogenetic position
Expanding the gnathostomata:

Source: http://tolweb.org/Gnathostomata/14843
(*) denotes extinct group
Note that we are classified with the lobe-finned fish, a group that includes coelacanths and lungfish. Expanding the sarcopterygii:

Source: http://tolweb.org/Sarcopterygii/14922
(*) denotes extinct group
(?) denotes uncertain phylogenetic position
Sarcopterygii expansion in more detail shows not only the tetrapoda (amphibia and reptilomorpha) but the significant stem-group taxa such as Ichthyostega, Ventastega and Acanthostega.
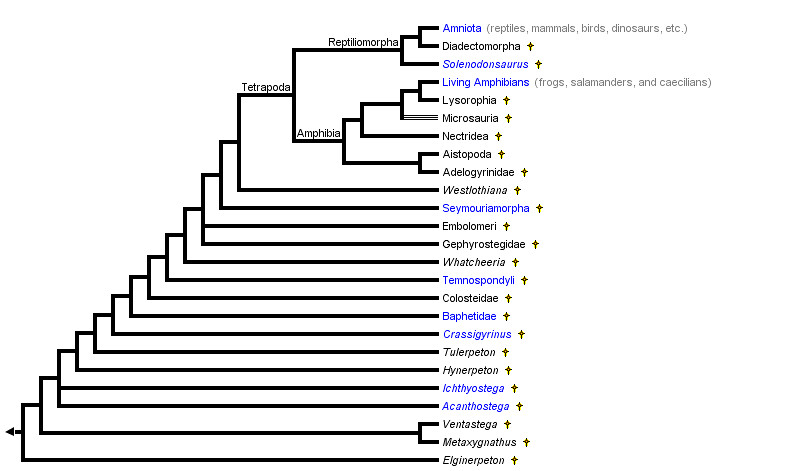
Source: http://tolweb.org/Terrestrial_Vertebrates/14952
This now puts us in a position to look at some of the significant fossil evidence.
Major Transitions from lobe-finned fish to tetrapods
Kenichthys: 395 million year old sarcopyterygian (lobe finned) fish from China. Its significance to evolution is the transitional nature of its nostrils. Tetrapods have a single pair of external nostrils that connect to the oral cavity via the choanae or 'internal nostrils.' Conversely, most living fish have two pairs of external nostrils on each side of their snout (anterior and posterior) that allow water to enter and exit the nasal cavity, respectively. Unlike tetrapods, the nasal cavity does not connect with the oral cavity. Figure 1 shows nostril positions on the heads of sarcopterygian fishes.
FIGURE 1. Nostril positions on the heads of sarcopterygian fishes.

a, reconstruction of Kenichthys campbelli head and cheek in lateral view. b, c, Youngolepis. d, e, Kenichthys. f, g, Eusthenopteron. b, d, f are lateral views; c, e, g are ventral views. In e, the (unknown) vomer is represented by its attachment area on the ethmoid. Not to scale. ch, choana; Dpl, dermopalatine; Enpt, entopterygoid; La, lacrimal; l.Ro, lateral rostral; Mx, maxilla; n.a, anterior nostril; n.p, posterior nostril; Pmx, premaxilla; Te, tectal; Vo, vomer.
Reprinted by permission from Macmillan Publishers Ltd: Nature 432, 94-97(4 November 2004) doi:10.1038/nature02843, copyright (2004)
Figures 2 and 3 show photographs of Kenichthys campbelli specimens and Kenichthys morphology, respectively:
FIGURE 2. Photographs of Kenichthys campbelli specimens.

a, b, V10493.61, ethmosphenoid in ventral [a] and anterior{b} views. c, d, V10493.60, ethmosphenoid in ventral [c] and lateral [d] views. e, V10493.58, ethmosphenoid in antero-lateral view. f, V10493.1, holotype, ethmosphenoid in antero-lateral view. g-h, V10493.101, ethmosphenoid in anterior view. i, V10493.81, lacrimal in external view. j, V10493.102, ethmosphenoid in anterior view. k, V10493.77, maxilla in external view. l, V10493.76, maxilla in external view. The green arrow indicates the free ventral margin of the lateral rostral; the red arrow in i indicates the slight 'step' marking the boundary of the tectal and prefrontal sutures. Scale bar, 2 mm.
Reprinted by permission from Macmillan Publishers Ltd: Nature 432, 94-97(4 November 2004) doi:10.1038/nature02843, copyright (2004)
FIGURE 3. Morphology of Kenichthys.

a–f, h, drawings of Kenichthys campbelli specimens. a, Restoration of ethmosphenoid in ventral view, based mainly on c and d. b, c, V10493.61, ethmosphenoid in anterior{b} and ventral [c] views. d, e, V10493.60, ethmosphenoid in ventral (d) and lateral (e) views. f, V10493.77, maxilla in external view. g, Maxilla of Medoevia (ref. 15), showing the diverging processes framing the choana. h, V10493.58, ethmosphenoid in antero-lateral view. Scale bar, 2 mm.
Reprinted by permission from Macmillan Publishers Ltd: Nature 432, 94-97(4 November 2004) doi:10.1038/nature02843, copyright (2004)
Two lines of evidence [3] suggested the posterior nostrils in fish became the choanae in tetrapods:
- Living lungfish have internal nostrils in their palate that resemble choanae
- Embryological data suggested that tetrapod choanae were homologous to the posterior nostrils of fish.
Fossil evidence from other lobe finned fish (specifically the osteolepiforms such as Eusthenopteron and Osteolepis) dating back to the Palaeozoic era has shown that they also had single external nostrils leading to openings in the palate similar to tetrapod choanae. Primitive lungfish (such as Diabolepis) however had two pairs of external nostrils. As Philippe Janvier points out:
Several theories have been proposed for the origin of the choanae of tetrapods and of their fossil osteolepiform relatives. The two main possibilities are that choanae are derived from the posterior external nostrils, but independently of development of the ‘internal nostrils’ of lungfishes, or that they are a novelty, and the posterior external nostrils have either disappeared or became the tear ducts. [4]One current scheme of tetrapod evolution shows two main 'sister' groups:
- Dipnomorphs (lungfish, Diabolepis and porolepiformes)
- Tetrapodomorphs (tetrapods and their closest fish relatives)

These depictions of the head and palate in some living and fossil lobe-finned fishes and tetrapods show the apertures of the nasal capsule — the anterior [a] and posterior [p] nostrils and the choanae [c]. The tree indicates the relationships of the groups, and the orange arrows indicate the two occurrences of the migration of the posterior nostril into the palate. In Kenichthys, the fossil studied by Zhu and Ahlberg, the posterior nostril forms a notch in the outer dental arcade, and is intermediate in position between that of an external nostril and that of choanae (p/c). The diagrams (1–3) on the right are of the left nasal cavity, in ventral view, in the three living forms. In lungfishes, the path of the maxillary nerve (mx) is medial to the posterior nostril; in tetrapods it is lateral to the choana. t, tear duct.
Reprinted by permission from Macmillan Publishers Ltd: Nature 432, 23-24(4 November 2004) doi:10.1038/432023a, copyright (2004)
The problem here is that while there was evidence of migration of nostrils in the Dipnomorph lineage, there was no comparable evidence in the tetrapodomorph lineage of posterior nostril migration. A 2004 paper in Nature on the posterior nostrils of the tetrapodomorph Kenichthys provides that evidence:
Here we present new material of Kenichthys, a 395-million-year-old fossil fish from China that provides direct evidence for the origin of the choana and establishes its homology: it is indeed a displaced posterior external nostril that, during a brief transitional stage illustrated by Kenichthys, separated the maxilla from the pre maxilla. [5]
As Janvier notes in his commentary:
Kenichthys is clearly a tetrapodomorph, and it displays exactly the intermediate condition that would be predicted when assuming that the tetrapod choanae were posterior nostrils displaced into the palate, in the same way as in lungfishes. The posterior nostrils (or already choanae?) of Kenichthys are actually right at the margin of the upper jaw, interrupting the outer dental ‘arcade’ formed by the premaxillary and maxillary bones. [6]
Kenichthys is therefore a tetrapodomorph with a nostril pattern that is clearly transitional.
Osteolepis: 390 million year old lobe-finned fish that has paired front and rear fins that has limb-like arrangement of the bones in its lobe fins, internal nostrils and amphibian-like skull and teeth. From an evolutionary point of view, its primitive middle ear is tetrapod-like in its configuration. This is a fish which has tetrapod features in its fins, skull and ears:
Osteolepis macrolepidotus fossil at the Museum für Naturkunde, Berlin. (Source: Wikipedia)
A specimen of Osteolepis macrolepidotus from the Middle Old Red Sandstone of Tynet Burn, Banffshire, in the Museum of the Zoological Department of University College, shows with perfect clearness a cast of the cavities left in the completely ossified otic capsule of both sides. The specimen is illustrated in the photograph.
The three semicircular canals, which automatically analyse any movement into three components referred to rectangular and rectilinear coordinates, form a theoretically perfect mechanism and have persisted without any serious modification through the vertebrates. [7]

Fig 1. Head of Osteolepis macrolepidotis, x 2 approx. (J Anat. 1925 July; 59(Pt 4): 385–386.)
Olf.C. olfactory capsule; A.C.S.A. ampulla of anterior vertical semicircular canal;
A.C.S.H. ampulla of horizontal canal; C.S.P. posterior vertical semicircular canal;
Hy. Md. Hyomandibular
In passing, this article was published in 1925, nearly ninety years ago. Creationists relying on relatively dated material from the second half of the last century may be excused for failing to comment on transitional fossils such as Tiktaalik roseae, but there is no excuse for failing to miss examples such as Osteolepis. Examples such as this demonstrate the poverty of special creationist research.
Eusthenopteron: 385 million year old lobe finned fish which has many transitional features indicating that it was well on the way to becoming a tetrapod.

Source: Wikipedia
- It has paired fins with distinct ulna, radius and humerus in the front fins, and fibula, tibia and femur in the rear fins. These are the arm and leg bones in tetrapods like us.
- It has internal nostrils
- It has teeth with infolded enamel such as seen in tetrapods

Source: Wikipedia
Jennifer Clack, one of the foremost experts on the fish to tetrapod evolutionary transition comments on Eusthenopteron, noting that while it has significant tetrapod-like features, it also has fish-like characteristics:
A feature shared by most fishes is the bone called the hyomandibula that supports the palate and controls movements of the gill apparatus. It is a long bone that pivots on the walls of the bone housing the brain and ear capsules, the braincase, and it has attachment points to the gill bars and the opercular series, the plate-like bones that cover and protect the gills. They are all part of the pumping mechanism used in breathing and feeding. Eusthenopteron was entirely typical of early fish in this respect. [8]

Source: http://palaeos.com/v....kVzf93uOlH.gif
{kind=link}

Source: http://palaeos.com/v....0Pq1JPw-RG.gif
{kind=link}
The presence of fin rays differentiates limbs from fins, so Eusthenopteron is technically classified as a fish. However, its fins have strong tetrapod-like features – forelimb bones homologous to the humerus, ulna and radius. It is a fish with limbs but no hands or feet. Creationists who talk about the creation of fixed ‘kinds’ in which limited evolution occurs do not have the luxury of resorting to this tactic, as objectively, this fossil has features that are fish-like, and features that are tetrapod-like. The presence of fin rays is one of the diagnostic criteria for defining a taxon as fish, so special creationists are fond of dismissing such fossils as fish with mosaic characteristics. However, that is exactly what palaeontologists regard this lobe-finned fish as being, namely a lobe-finned fish with a mosaic of tetrapod and fish characteristics is a fossil with transitional features.

Source: http://palaeos.com/v...nPectFinAll.gif
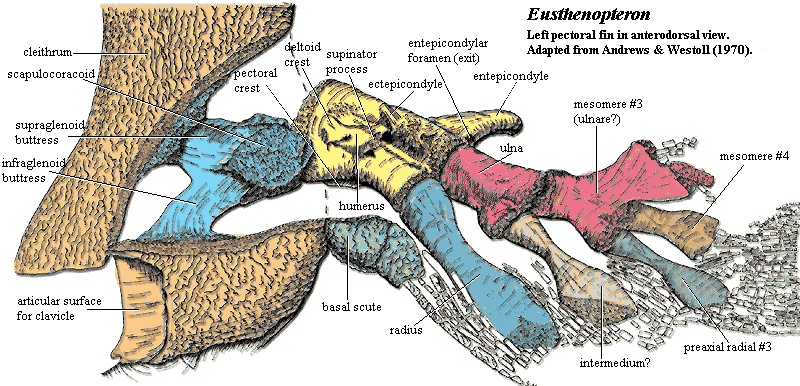{kind=link}
Gogonasus: this is a lobe finned fish that has a tetrapod-like middle ear and bones in its pectoral fins that were tetrapod-like. Its skeletal anatomy [9] shows that it was more tetrapod-like than Eusthenopteron.
FIGURE 1. Cranial features of Gogonasus andrewsae, NMV P221807.

a, Skull in lateral view. b, c, skull in dorsal view. d, e, X-ray micro-tomograms showing the spiracular region in lateral (d) and anterior (e) views. f–h, Restorations of palatoquadrates showing the spiracular chamber (shaded) for Panderichthys (f), Gogonasus (g) and Eusthenopteron (h); panels f and h are based on ref. 9. ET, extratemporal bone; PO, postorbital; PP, postparietal; Spir, spiracular opening; Sp. lam., down-turned lamina of tabular for spiracular chamber; Sq, squamosal; ST, supratemporal; Ta, tabular. All bones whitened with ammonium chloride. - Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202
FIGURE 2. Pectoral fin skeleton of Gogonasus andrewsae, NMV P221807.

a–d, Right humerus in dorsal (a), mesial (b), proximal (c) and distal (d) views. e, Right ulnare in ventral view. f, Left humerus, ulna and radius articulated in dorsal view. g, Reconstructed left pectoral fin in dorsolateral view. h, Left pectoral distal elements. i, Key to h. A4, A5, distal mesomeres of the pectoral fin; Cap. hum., caput humeri; Ent. for., entepicondylar foramen; Ent. pr., entepicondyle; Pr. ven., ventral process on entepicondyle; Rad., presumed radial element; Rad. art., radial articulation; Rad. pr., radial process; Ul. pr., postaxial process of ulnare. All (except g) whitened with ammonium chloride. Scale bar, 1 cm for a–d, f–i; 0.5 cm for e. - Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202
- Loss of all midline fins
- Flatter skull with a longer snout and larger, more dorsally placed eye sockets.
- Humerus and shoulder girdle larger and more like that of a tetrapod as compared to Eusthenopteron.
- Spiracular cleft is larger than that in Eusthenopteron, and approximates that of a tetrapod. In fact, it may have been open to the outside, which is a feature associated with air breathing.
- The hyomandibular bone (homologous to the stapes, one of the middle ear bones in tetrapods) is shorter than Eusthenopteron, but still connected to the opercular bones.

Reconstruction of Panderichthys. Source: http://devonianlife....erichthys 4.jpg
{kind=link}
Figure 1 Pictures and drawings of Panderichthys rhombolepis, specimen GIT434-1.

a, Outline of the body of Panderichthys. Grey shading indicates preserved portions of Panderichthys rhombolepis specimen GIT 434-1. Redrawn from ref. 14. b, Panderichthys rhombolepis specimen GIT 434-1 with head (h) and body (b) outlined. The pelvic girdle and fin are shaded in orange. c, Pelvic girdle and fin. The matrix is distinguished from the fossil by an overlay of grey shading. d, Specimen drawing. F, femur; Fi, fibula; Fre, fibulare; Int, intermedium (proximal end of the); Pel, pelvic girdle; T, tibia. Vertical hatching indicates broken bone; grey shading indicates matrix; circles indicate thin dermal bone covering. e, Reconstruction of the pelvic fin. Thick outline indicates preserved margin, thin outline indicates inferred margin, dotted lines indicate uncertain margin. Solid black scale bars, 10 mm. - Boisvert CA "The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion" Nature (2005) 438:1145-1147
One researcher points out that the anatomical details of Panderichthys shed considerable light on the evolution of tetrapod locomotion:
The morphology of the pelvic fin of Panderichthys is a combination of primitive, transitional and unique characteristics that provide crucial information about the evolution of the tetrapod hindlimb and the origins of tetrapod locomotion.
FIGURE 2. Comparison of pectoral and pelvic fins.
The pectoral fin is more tetrapod-like than the pelvic fin in that the shape of the humerus is intermediate between those of osteolepiforms and basal tetrapods. [11]

Pectoral (a, c, e) and pelvic fins (b, d, f) of Eusthenopteron (a, b), Panderichthys (c, d) and Acanthostega (e, f) all in ventral view. F, femur; Fi, fibula; Fre, fibulare; H, humerus; Int, intermedium; R, radius; T, tibia; U, ulna; Ure, ulnare. Thick outline indicates preserved margin; thin outline indicates inferred margin; dotted lines indicate uncertain margin. Scale bars, 10 mm. Panels a and b are redrawn from ref. 3, c is from ref. 11, scale bar is from ref. 5, and e and f are redrawn from ref. 6. - Boisvert CA "The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion" Nature (2005) 438:1145-1147
Elpistostege: late Devonian lobe-finned fish dated around 385-380 million years ago that differs from Panderichthys in having longer snout and smaller, more rounded orbits. It resembles tetrapods in having:
- Dorsoventrally flattened skulls
- Supraorbital ridges or “eyebrows”
- External nostrils close to the jaw margin

Source: http://www.miguasha....stege-fmd_g.jpg
{kind=link}

Source: http://fossils.valdo...ls_dev/f17.html
Per Ahlberg, one of the leading experts in early tetrapod evolution notes that this fossil is a grade intermediate [12] between Panderichthys and tetrapods.
Tiktaalik: late Devonian fish [13] which is similar to Panderichthys but is even more like tetrapods, to the point that is unofficially referred to as a ‘fishapod’:

Source: Wikipedia
- Longer snout with larger eyes
- Shoulder girdle not in contact with head. This allows greater head mobility to permit raising and lowering with far more ease than other fish
- Hyomandibular bone is shorter than in Panderichthys indicating changes to the gill region and breathing mechanism
- Spiracular chamber opening is more rounded.

a, Left lateral view; b, dorsal view with enlargement of scales; and c, ventral view with enlargement of anterior ribs. See Fig. 3 for labelled drawing of skull in dorsal view. Abbreviations: an, anocleithrum; bb, basibranchial; co, coracoid; clav, clavicle; clth, cleithrum; cbr, ceratobranchial; ent, entopterygoid; hu, humerus; lep, lepidotrichia; mand, mandible; nar, naris; or, orbit; psp, parasphenoid; ra, radius; suc, supracleithrum; ul, ulna; uln, ulnare. Scale bar equals 5 cm. - Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature. 2006; 440(7085):757-63
a, Dorsal view of body with scales removed depicting orientation of ribs as preserved in NUFV 108. b, Lateral view. The ribs are shown in dorsoventral orientation. Number of ribs is estimated from the incompletely preserved series in NUFV 108. - Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature. 2006; 440(7085):757-63
The most important feature of Tiktaalik is that it has a wrist, which takes its upper fin even closer to being classed as a limb:
The array of joints in the distal fin is functionally similar to the multiple transverse joints that characterise the carpal, metacarpophalangeal and interphalangeal joints of the tetrapod manus. The distal endoskeleton of Tiktaalik invites direct comparisons to the wrists and digits of limbed vertebrates. The intermedium and ulnare of Tiktaalik have homologues to eponymous wrist bones of tetrapods with which they share similar positions and articular relations. In both Tiktaalik and early tetrapods, the ulnare is block-shaped and articulates with multiple radials or digits, whereas the intermedium is a simple rod. The formation of a mobile transverse joint at the distal margin of these bones in Tiktaalik presages the establishment of a functional proximal carpal joint.”
“As in the digits and phalanges in a tetrapod limb, the inter-radial joints distal to this primordial wrist are more or less transversely aligned and capable of flexion and extension. The occurrence of multiple distally facing radial rows that are capable of flexion and extension is a likely antecedent condition to the dactyly of early tetrapods. The transformation of fins to limbs, then, probably entailed the elaboration and proliferation of structures, joints and functions already present in the fins of fish such as Tiktaalik.” [14]

a, Stereo pair of left pectoral fin of NUFV 108 in dorsal view showing disparity in size and position of anterior (alp) and posterior (plp) unjointed lepidotrichia and the relative position of dermal girdle elements. b, Right pectoral fin of NUFV 110 in anterior view showing preservation of anterior lepidotrichia (alp), clavicle (cl), scales (sc) and endochondral bones in articulation (H, humerus; U, ulna; u, ulnare; r, radials). The anterior lepidotrichia terminate at the elbow, thus allowing a full range of flexion at that joint. c, Right pectoral fin of NUFV 110 in ventral view showing positions of coracoid (co) and endochondral and dermal fin elements. an, anocleithrum; cb, ceratobranchial; clth, cleithrum; int, intermedium; ri, rib; suc, supracleithrum. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006 440(7085):764-71.

a, Dorsal view; b, ventral view. Elements with stipple shading were preserved in articulation in NUFV 109 and prepared in the round. Elements with a dashed outline are reconstructed based on their presence in the articulated distal fin of NUFV 110. It is not known how many radials lie distal to the first, second and fourth in the proximal series. Note the dorsal expansion of the distal articular facets on the ulnare and third distal radial/mesomere. The dorsal expansion of these facets would have facilitated extension of the distal fin. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006 440(7085):764-71.

a, Dorsal view; b, ventral view; c, medial view; d, posterior view. ac gle, anterior cam of glenoid facet; cf, coracoid foramen; clth, cleithrum; co, coracoid; gle, glenoid facet; sp, scapular process. The hatched area is covered by matrix. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006 440(7085):764-71.

Unlike other tetrapodomorph fishes (1), Tiktaalik has reduced the unjointed lepidotrichia, expanded the radials to a proximal, intermediate and distal series, and established multiple transverse joints in the distal fin. The fin also retains a mosaic of features seen in basal taxa. The central axis of enlarged endochondral bones is a pattern found in basal sarcopterygians and accords with hypotheses that a primitive fin axis is homologous to autopodial bones of the tetrapod limb. In some features, Tiktaalik is similar to rhizodontids such as Sauripterus. These similarities, which are probably homoplastic, include the shape and number of radial articulations on the ulnare, the presence of extensive and branched endochondral radials, and the retention of unjointed lepidotrichia. Figures redrawn and modified from Glyptolepis, Eusthenopteron, Panderichthys, Acanthostega and Tulerpeton. - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.

a, b, Anterolateral view. c, d, Ventral view. a, c, Resting posture with the fin partially flexed at the antebrachium. In this position the radius is slightly more flexed than the ulna. b, d, Resistant contact with a firm substrate entails flexion at proximal joints and extension at distal ones. The shoulder joint is flexed by ventral muscles, including the trans-coracoid muscle. The elbow is flexed (d, arrow 1), with slight pronation of the radius (d, arrow 2) and rotation of the ulna (d, arrow 3). The transverse joints distal to the ulnare and intermedium are extended (d, arrows 4). - Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440(7085):764-71.
Any question as to the transitional status of this fossil is explicitly put to bed by the authors who note:
The pectoral skeleton of Tiktaalik is transitional between fish fins and tetrapod limbs. Comparison of the fin with those of related fish reveals that the manus is not a de novo novelty of tetrapods; rather, it was assembled in fishes over evolutionary time to meet the diverse challenges of life in the marshes of Devonian aquatic evosystems. [15]
To summarise the above comments from the research paper:
- The bones in the pectoral fin of Tiktaalik perform similar functions to those in the tetrapod upper limb
- Furthermore, these bones are homologous to those in the tetrapod limb
- Although Tiktaalik is a fish by definition (it has scales and the presence of rays means it has fins instead of limbs), structurally and functionally, it is clearly transitional between fish and tetrapod to the point that it blurs the transition and fully warrants the name ‘fishapod’
The original papers were silent on the pelvic girdle of Tiktaalik; last year, Shubin et al published an analysis of the its pelvic girdle which apart from providing the final major piece in the anatomical puzzle also provided further details on its transitional status, as well as information on the evolution of tetrapod locomotion:
The earliest tetrapods have robust limbs, particularly hind limbs that are enlarged and supported by a number of mod- ifications to the pelvic girdle. In contrast, the closest relatives of tetrapods maintain small and weakly ossified pelvic appen- dages as compared with the pectorals. This observation has led to the “front wheel drive” hypothesis that held that the closest relatives of tetrapods emphasized pectoral support and locomotion whereas significant pelvic support and locomotion was a tetrapod innovation. The discovery of pelvic girdle and fin material of the tetrapodomorph Tiktaalik roseae reveals a tran- sitional stage in the origin of the pelvic girdle and appendage: although retaining primitive skeletal architecture, these ele- ments are enhanced in size and robusticity much like tetrapods. [16]

Type specimen (NUFV108): ventral surface of cranial block (figured in ref. 6) aligned in preserved position with ventral view of the block containing the pelvic fin. (Inset) Line diagram of lepidotrichia and preserved portions of endochondral bones of pelvic fin. f, fin; i, intermedium?; l, lepidotrichia; r, radials.

Acanthostega (Top), Tiktaalik (Middle), and Eusthenopteron (Bottom) reconstructions with pectoral and pelvic girdles.
Ventastega: this is a late Devonian tetrapod with fish-like features. [17] It is classified as a tetrapod on the basis of material from its lower jaw and girdle.

Ventastega reconstruction. Source: Wikipedia
Of note are:



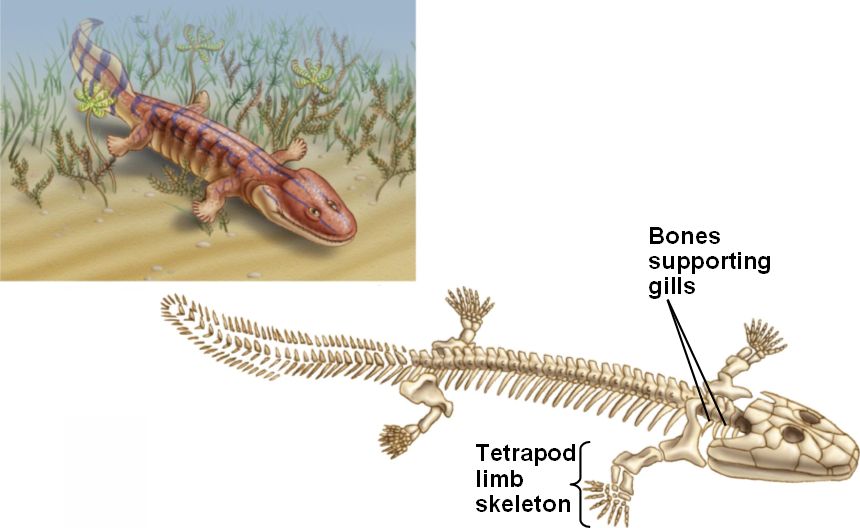
Acanthostega: was a Devonian tetrapod living around 370-360 million years ago. It had limbs with digits, but retained a number of fish-like characteristics. Despite the presence of legs, it was poorly suited to terrestrial life as it could not support its weight on land, and was therefore mainly aquatic.

Ventastega: this is a late Devonian tetrapod with fish-like features. [17] It is classified as a tetrapod on the basis of material from its lower jaw and girdle.
Ventastega reconstruction. Source: Wikipedia
Of note are:
- Its shoulder and hip girdle, which closely resemble that of the tetrapod Acanthostega, allowing one to infer the presence of digits.
- Its skull, which is intermediate in appearance between that of the fish Tiktaalik and the tetrapod Acanthostega.
- Its braincase, which shows a mix of fish-like and tetrapod-like features.
a, b, Associated skull roof (LDM G 81/775) and cheek (LDM G 81/776) in dorsal (a) and left lateral (b) views, anterior at the top. The internasal fontanelle, orbit and spiracle are indicated in a. c, d, The same specimen without the cheek in left lateral (c) and ventral (d) views, anterior at the top, showing the three-dimensionally preserved braincase. The parasphenoid and sphenoid are indicated. e, Complete lower jaw (LDM G 81/777) in medial view with coronoid fangs shown. Scale bar, 10 mm. 'LDM G' denotes the geology collections of Latvijas Dabas Muzejs, the Natural History Museum of Latvia. - Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". Nature (2008) 453:1199-1204
a, Right anocleithrum (LDM G 81/778) in lateral and anterior views (from left to right), showing overlap for cleithrum. b, Left cleithrum (LDM G 81/779) and partial scapulocoracoid in lateral, anterior and mesial views (from left to right). Note the broad shallow subscapular fossa (s.fossa) and the partially preserved glenoid canal. c, Interclavicle (LDM G 81/601) in ventral view showing clavicular overlaps. d, A probable tetrapod rib (LDM G 81/781). e, Right ilium (LDM G 81/780) in anterior, lateral, mesial and dorsal views (from left to right), showing the iliac neck, dorsal process and posterior process. f, A probable tetrapod caudal fin lepidotrichium (LDM G 81/782) on a block of matrix. Scale bar, 10 mm; all specimens shown to same scale. - Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". Nature (2008) 453:1199-1204
a, Whole-body reconstruction showing known skeletal elements on a body outline based on Acanthostega (modified from ref. 5; original Acanthostega body reconstruction by M. I. Coates). Scale bar, 10 cm. b, c, Skull reconstruction in lateral and dorsal views, based on material presented here and described previously21. d, Reconstructed association of skull and shoulder girdle in lateral view. e, Shoulder girdle in anterior view. Curvature of cleithrum based on LDM G 81/522 (ref. 21). Unknown bones are indicated with vertical hatching. Scale bar for b–e, 10 mm. f, g, Life reconstructions of head in lateral and dorsal views (copyright P. Renne, 2007). an, anocleithrum; ang, angular; cla, clavicle; clei, cleithrum; de, dentary; fr, frontal; icl, interclavicle; i.fon, internasal fontanelle; it, intertemporal; ju, jugal; la, lacrimal; mx, maxilla; m.ro, median rostral; na, nasal; pa, parietal; pmx, premaxilla; po, postorbital; pof, postfrontal; pop, preopercular; pospl, postsplenial; pp, postparietal; prf, prefrontal; pter, pterygoid; qj, quadratojugal; sang, surangular; scapcor, scapulocoracoid; spl, splenial; sq, squamosal; ta, tabular. - Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". Nature (2008) 453:1199-1204
Acanthostega: was a Devonian tetrapod living around 370-360 million years ago. It had limbs with digits, but retained a number of fish-like characteristics. Despite the presence of legs, it was poorly suited to terrestrial life as it could not support its weight on land, and was therefore mainly aquatic.
Acanthostega reconstruction. Source: Wikipedia
Fish-like features included:

Source: http://www.theclacks...canthostega.jpg

Source: http://www.nicerweb....canthostega.jpg
Fish-like features included:
- Gill bars
- No true elbows, knees, wrists or ankles
- Long fin rays around tail
Source: http://www.theclacks...canthostega.jpg
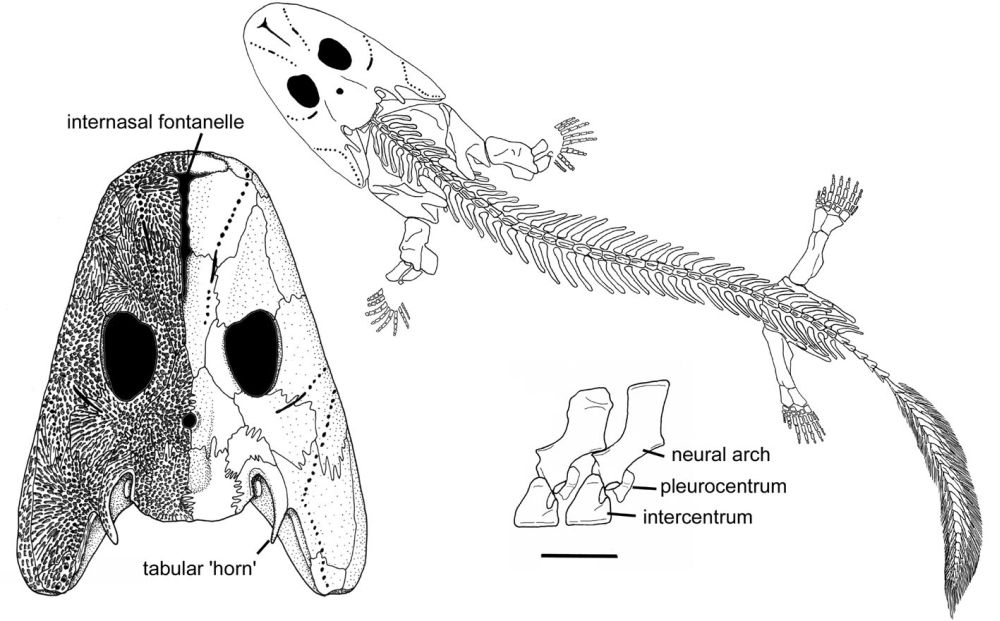{kind=link}
Source: http://www.nicerweb....canthostega.jpg
{kind=link}
Of course it was a tetrapod as it had four limbs with digits. Other specific tetrapod features [18] included:
- Head not joined to shoulder girdle
- Ear bone fitted into braincase fenestra
- Large pelvic girdle and sacral rib
- Femur and humerus were of similar size
- Tibia and fibula attached to ankle bones
- Digits on each limb

Pectoral endoskeleton of tetrapodomorph fish and a tetrapod in ventral view. Pectoral fins of: a, Eusthenopteron; b, Panderichthys; and c, Tiktaalik. d, Limb of Acanthostega. H, humerus; Int, intermedium; R, radius; U, ulna; Ure, ulnare. Scale bar, 1 cm. - Nature (2008) 456:636-638

Source: http://palaeos.com/v....kmzIKtTAjT.gif
{kind=link}
Clack is worth quoting at length to show why Acanthostega is regarded as an excellent transitional fossil:
The fish-like features of Acanthostega include a tail fin supported by long bony rays, to make an oar shape. Fin rays had been found in the tail of Ichthyostega, but they were much shorter and less numerous. The ribs of Acanthostega are short and slender and much the same all down the length of its body… Ichthyostega was known to have had broad overlapping ribs at least in its trunk region. In the skull, Acanthostega has a slot running down the midline of the snout, similar to the gap found there in Ventastega...Acanthostega also retains a set of well-ossified grooved gill bars, suggesting that these were still actively involved in gill breathing, though it probably breathed air by gulping as well.In other respects, Acanthostega showed some typical tetrapod features. It has a robust pelvic girdle and a large femur. It is the pelvic region and the hind limb that most conspicuously distinguish a fish from a tetrapod, and that distinction is still valid among modern animals. In fish, the pelvic girdle is usually small and not attached to the vertebral column and the pelvic fin is diminutive, whereas in tetrapods, the pelvic girdle is large, attached to the vertebral column and bears the substantial hind limbs that provide most of the propulsive power in walking. In Acanthostega, although the pelvic girdle is enlarged, it is still relatively small compared with later tetrapods and was only attached to the column by soft tissue rather than a bony junction.Some excellent specimens showed that it had eight digits on the forearm…and almost certainly eight (or maybe more) on the hind limb. That they are so neatly laid out in the fossil may be because they were enveloped in a web of skin. The wrist bones were not ossified, but it is clear that the digits were arranged in a broad arc around where those bones would have been, but the result would not have made a flexible or supportive wrist. The limb was in effect a paddle…Similarly, although the ankle bones are well ossified, they do not show an obvious joint surface along which the limb could bend. Again, the limb seems to have been a paddle. From that evidence, it was suggested that limbs with digits first evolved not for walking on land but for swimming or wading through water.In the braincase and ear region, Acanthostega showed features in which it was distinctively more tetrapod-like than fish-like. In tetrapods, rather than a hyomandibula pivoting on the braincase wall, essentially the same bone but now termed the stapes, fits into a hole in the braincase wall called the fenestra vestibuli or fenestra ovalis. Acanthostega shows this pattern in a very early form. The arrangement later became modified into the combination of a stapes and a fenestra ovalis that is characteristic of all tetrapods and is still part of the hearing mechanism in humans. [19]

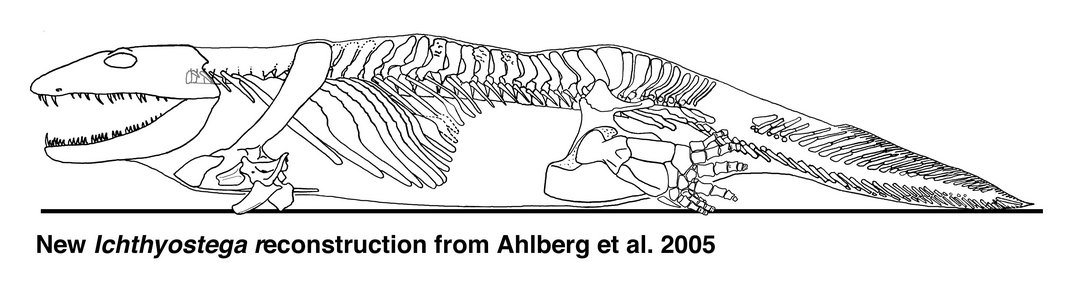
Ichthyostega reconstruction. Source: Wikipedia

Source: http://tolweb.org/tr...ostega.300a.jpg (Reconstruction derived from research by Per Ahlberg)
{kind=link}
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. Copyright © 2005 Jennifer A. Clack

Reconstructions of the skull of Ichthyostega in dorsal (top left), ventral (top right) and lateral views (bottom). © 1997 J. A. Clack.
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
Copyright © 2005 Jennifer A. Clack

Skull roof of Ichthyostega. MGUH VP 6064 Geological Museum, Copenhagen. Photo J. A. Clack. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...kull_fossil.JPG)
Clack again summarises its status as a key transitional fossil:

Hindlimb of Ichthyostega; specimen collected in 1987. Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...a_hind_limb.JPG)

Photographs of part and counterpart superimposed to show seven digits (specimen collected in 1987). Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...ostega_foot.JPG)

Source: Wikipedia


{kind=link}
Clack again summarises its status as a key transitional fossil:
If Acanthostega is a mixture of fish-like and tetrapod-like characters, Ichthyostega is a mixture of apparently aquatic specializations with others that appear more modified for land excursions. Recent study has revealed a rather different animal from the old image of an overlarge salamander-like body form.
One of the parts of the anatomy that had puzzled earlier workers was the ear region. Using newly collected and prepared fossil material, together with microcomputed tomography scanning of key fossils, the unique arrangement found in Ichthyostega has been interpreted as a highly specialized underwater auditory organ…By contrast, work on its postcranial skeleton has suggested that it may have had a unique form of locomotion on land. The vertebral column is differentiated into separate regions, including a lumbar portion that seems to have accommodated dorsoventral flexion The shoulder girdle and forearm are large and robust, suggesting extensive musculature, perhaps for pulling the animal along on land. Its hind limb is quite similar in construction to that of Acanthostega, that is to say a paddle with no obvious ankle joint, and it has seven toes arranged in a unique pattern. A group of three small toes form a strengthening bar along the leading edge, with four stouter ones behind. It is likely that, as in Acanthostega, these were contained in a web of skin. The hind limb may have given stability and purchase on the substrate and been used as a flipper in water, but it was not a conventional walking leg. It is no surprise that this peculiar morphology caused problems for early workers trying to understand Devonian tetrapods. It shows that even by that time, tetrapods had diversified into a wider range of morphologies than was previously appreciated. Analysis of all the skull specimens of Ichthyostega has shown changes to proportions and skull bone ornamentation that varies throughout its history and that document microevolution at the specific level in these very early tetrapods. Skulls from lower formations are narrower, with finer ornament, than those from higher up in the geological sequence. [20]
Hindlimb of Ichthyostega; specimen collected in 1987. Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...a_hind_limb.JPG)
{kind=link}
Photographs of part and counterpart superimposed to show seven digits (specimen collected in 1987). Geological Museum, Copenhagen MGUH Field number 1349. Photo M. Coates. Copyright © 1997 University Museum of Zoology, Cambridge. (http://tolweb.org/tr...ostega_foot.JPG)
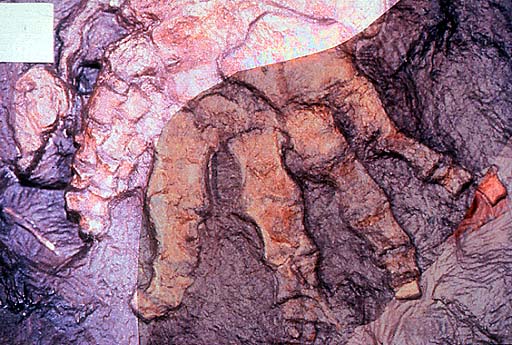{kind=link}
Pederpes: this was a tetrapod living around 360 to 350 million years ago that had five digits on its hind limbs, and six digits – five formed and one vestigial – on its front limbs. This details more evolutionary change from the earlier tetrapods as inferred from Ichthyostega and Acanthostega, which had more than five digits.
Source: Wikipedia
“This animal, belonging to a group known as the whatcheeriids, not only shows some primitive features in which it resembles the Devonian forms but also has more advanced features that suggest the beginnings of terrestriality. Its hind limb was fairly typical of a conventional tetrapod from the Late Carboniferous and appears to have had five digits. The foot appears to have pointed forward as in a terrestrially walking animal, rather than to the side as in Acanthostega. However, the forelimb may have had more than five digits, because the only two that are known are both extremely small and resemble the supernumeraries of Acanthostega and Ichthyostega. Pederpes has broad flanges on its ribs that resemble those of Ichthyostega in some ways. Its stapes is like that of Acanthostega.” [21]
a, GLAHMS 100815 Pederpes finneyae, photograph of holotype and only specimen. Scale bar, 100 mm; b, Map of the holotype, showing elements preserved on the reverse in grey. Scale bar, 10 mm. Fro, frontal; intemp, intertemporal; jug, jugal; lac, lacrimal; max, maxilla; nas, nasal; pmx, premaxilla; po, postorbital; pofr, postfrontal; prefr, prefrontal; preop, preopercular; qj, quadratojugal; scapcor, scapulocoracoid. - Clack J.A. "An early tetrapod from 'Romer's Gap'" Nature (2002) 418:72-76 doi:10.1038/nature00824
Close-up of the skull of Pederpes showing the stapes. - Clack J.A. "An early tetrapod from 'Romer's Gap'" Nature (2002) 418:72-76 doi:10.1038/nature00824

References
a, Humerus, radius, ulna and manual digits from the right forelimb. The minute irregular elements at the base of the tiny digit might be remains of metacarpals. b, Pedal digits and femur from the right hindlimb, shown reversed. c, Reconstructions of manual (left) and pedal digits (right) of Pederpes to the same scale, showing tiny manual digit and asymmetrical metatarsals respectively. Scale bar, 10 mm. In Pederpes, the phalanges of the tiny manual digits are about one-third the length of the pedal phalanges, whereas in Acanthostega and the Carboniferous forms such as Greererpeton and Proterogyrinus, manual phalanges are about one-half the length of the pedal phalanges. - Clack J.A. "An early tetrapod from 'Romer's Gap'" Nature (2002) 418:72-76 doi:10.1038/nature00824
Again, we see the mosaic of fish-like and tetrapod-like features, but with a definite progression towards terrestrial life. This – yet again – is another transitional fossil which along with the earlier taxa shown illustrate large-scale evolutionary change from lobe-finned fish to tetrapods.
Conclusion
One of the classic tropes of evolution is that of fish evolving limbs. It is also the subject of much special creationist misrepresentation. As we've seen, the evidence for the origins of tetrapods from lobe finned fish is robust, and is a classic example of large-scale evolutionary change.
References
1. Hunt K Transitional Vertebrate Fossils FAQ
2. Material in this post is taken with permission from this article.
3. Janvier, P. "Wandering nostrils." Nature (2004) 432:23-24
4. ibid, p 23-24
5. Zhu, M. & Ahlbert, P. E. "The origin of the internal nostril of tetrapods"Nature (2004) 432:94-97
6. Janvier op cit p 24
5. Zhu, M. & Ahlbert, P. E. "The origin of the internal nostril of tetrapods"Nature (2004) 432:94-97
6. Janvier op cit p 24
7. Watson DMS “The Internal Ear of Osteolepis” J Anat (1925) 59:385-88
8. Clack J.A. “The Fish-Tetrapod Transition: New Fossils and Interpretations” (2009) Evo Edu Outreach 2:213-223
9. Long JA et al "An exceptional Devonian fish from Australia sheds light on tetrapod origins" Nature (2006) 444:199-202
10. Boisvert C.A. The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion. Nature. (2005) 438:1145-7.
11. ibid, p 1145
12. Ahlberg P.E. et al “A near-tetrapod from the Baltic Middle Devonian” Palaeontology (2000) 43:533-548
10. Boisvert C.A. The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion. Nature. (2005) 438:1145-7.
11. ibid, p 1145
12. Ahlberg P.E. et al “A near-tetrapod from the Baltic Middle Devonian” Palaeontology (2000) 43:533-548
13. Daeschler E.B., Shubin N.H, Jenkins F. “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature(2006) 440:757-63
14. Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. (2006) 440:764-71.
15. Shubin, Daeschler, Jenkins. op cit, p 769
15. Shubin, Daeschler, Jenkins. op cit, p 769
16. Shubin N.H., Daeschler E.B. Jenkins F.A. "Pelvic girdle and fin of Tiktaalik roseae" Proc Natl Acad Sci USA (2014) 111:893-899
17. Ahlberg, PE. Clack JA, Luksevics E, Blom H and Zupins I "Ventastega curonica and the origin of tetrapod morphology". Nature (2008) 453:1199-1204
18. Clack JA. Acanthostega. A very primitive tetrapod from the Late Devonian with eight fingers and toes. A 'transitional form' between 'fishes' and 'tetrapods'. Accessed: 6th January 201219. Clack J.A. (2009) p 219
20. op cit p 219-220
21. op cit p 220